







































B cells
B cells are lymphocytes (WBCs) that participate in humoral immunity by producing antibodies in response to antigen stimulation.
▼ activation : B-1 : B-2 : BCRs : CDRs : granzymes : helper T cells : life-span B cells : lymphopoiesis : memory B : naïve B cells : NK cells : NK receptors : NK cells attack viral infected cells : perforin : plasma B : stimulation : surface-immunoglobulins : surface receptors : VDJ recombination ▼
Surface membrane-associated immunoglobulins (IgD and IgM) act as B cell receptors (BCRs), and the enormous variety of antigen recognition sites is attributable to VDJ recombination (alternative splicing) of peptide sequences encoded by V, D, and J genes. The variable region of immunoglobulins includes the recognition sites or complementarity determining regions (CDRs).
Lymphopoiesis, which takes place in the bone marrow of almost all mammals, produces small lymphocytes, large granular lymphocytes (NK) cells, B lymphocytes (precursors of plasma cells, T lymphocytes, and lymphoid dendritic cell. Recognition of self during lymphopoiesis permits anergy (suppression of self-attack).
Naïve B cells each have one of millions of distinct surface antigen-specific receptors, yet have not encountered their specific, cognate antigen. With a life-span of only a few days, many B cells die without ever encountering their cognate antigen. Naïve B cells are activated when the BCR binds to its cognate antigen. This antigen-Ig binding must be coupled with a signal from a helper T cell in order to activate the B cell.
Once activated, B lymphocytes:
● differentiate into one of the B cell types (directly or through intermediate, germinal center reactions)
● plasma cells produce antibodies against the antigenic stimulus, or memory cells are primed for subsequent activation by the antigen
Types of B cell:
● B-1
● B-2
● Plasma B cells
● Memory B cells
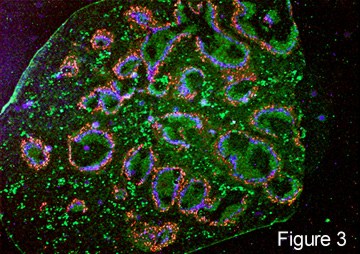
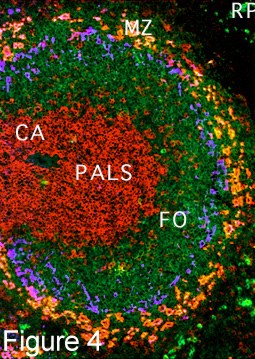
After newly formed B cells exit generative sites in fetal liver or adult bone marrow they undergo selection events that may involve interactions with self or with external antigens. Selective events can influence the phenotype and functional characteristics of B cells. B cell receptor-mediated events also influence lymphoid organs localization as marginal zone B cells in the spleen, as follicular (B-2 cells), as well as B-1 cells in the peritoneal and pleural cavities. [] fluorescence micrograph spleen, fm high power in which T cells form periarteriolar lymphocyte sheath (PALS) (red) and B-2 cell follicles (green) []
B-1 cells are the first B cells produced in the fetus, and in adults are located primarily in the peritoneal and pleural cavities. B1 cells are believed to operate in the innate response to infection by viruses and bacteria, and usually show preferential responses to T cell-independent antigens. The diversity of B-1 lymphocytes is attributed to their recombinatorial recombination, in which there is a preferential recombination between D-proximal VH gene segments. B-1 lymphocytes express (polyspecific) IgM in greater quantities than they express IgG, and the ability of B1 cells to respond to isotype switch commitment factors such as interleukin-4 may be secondary to their production of IgM. B-1 cells express CD5, which binds to CD72 to mediate B cell-B cell interactions.
B-2 cells are conventional B lymphocytes that are produced postnatally (unlike fetal B-1 cells) and are replaced from the bone marrow.
Plasma B lymphocytes are committed to production of copious amounts of monoclonal antibodies.
Memory B lymphocytes are long-lived, stimulated B lymphocytes that are primed for rapid response to a repeated exposure of the priming antigen. Memory B cells are generated in lymphoid tissue after B cell activation/proliferation and reside in the bone marrow, lymph nodes, and spleen. High affinity surface immunoglobulins enable their activation by lower levels of cognate antigen than are naïve B cells.
NK cells are differentiated from killer T cells. NK, natural killer cells constitute a corps of circulating lymphocytes that are constitutively specialized to attack cancerous cells and virus infected cells. Preprogramming for target recognition, coupled with the absense of need for backup by a clone of identical cells, renders NK cells capable of rapid (innate) response to pathogens. NK attack involves the exocytosis of cytoplasmic granules containing perforin and granzymes. Perforin forms pores in the plasma membrane of attacked cells through which serine-protease granzymes enter, cleaving caspase precursors and triggering apoptosis.
Individuals inherit multiple, polymorphic genes for NK receptors, so the assemblage of NK receptors differs between individuals. NK cells carry two forms of surface receptors:
● killer inhibitory receptors (KIRs) transmit an inhibitory signal when they encounter class I MHC molecules on a cell surface. (By contrast, T cells only recognize antigens that are presented by a MHC molecule.)
● activating receptors, which activate the NK cell upon binding to a target cell
Viral infection often causes suppression of MHC expresion, leading to a reduction of inhibition of NKs by its killer inhibitory receptors. This double negative renders the virus infected cell a target for killing by NK cells.
"About 85% of peripheral B cells are phenotypically mature and display first-order exponential kinetics defined by a half-life of 5-6 weeks, whilst the remainder are short-lived with a life span of several days."[s]
[] tem plasma cell [] micrograph macrophage surrounded by normal plasma cells [] micrograph macrophage & plasma cells []
▼ activation : BCRs : CDRs : helper T cells : life-span B cells : lymphopoiesis : naïve B cells : surface-immunoglobulins : surface receptors : VDJ recombination ▼
Tables Fc receptors Immune Cytokines Immunoglobulins Cell Adhesion Molecules Cell signaling Receptor Tyrosine Kinases (RTKs) Receptor Signal Transduction Second Messengers
▲ Top ▲
tags [Immunology][lymphocyte]

▼ activation : B-1 : B-2 : BCRs : CDRs : granzymes : helper T cells : life-span B cells : lymphopoiesis : memory B : naïve B cells : NK cells : NK receptors : NK cells attack viral infected cells : perforin : plasma B : stimulation : surface-immunoglobulins : surface receptors : VDJ recombination ▼
Surface membrane-associated immunoglobulins (IgD and IgM) act as B cell receptors (BCRs), and the enormous variety of antigen recognition sites is attributable to VDJ recombination (alternative splicing) of peptide sequences encoded by V, D, and J genes. The variable region of immunoglobulins includes the recognition sites or complementarity determining regions (CDRs).
Lymphopoiesis, which takes place in the bone marrow of almost all mammals, produces small lymphocytes, large granular lymphocytes (NK) cells, B lymphocytes (precursors of plasma cells, T lymphocytes, and lymphoid dendritic cell. Recognition of self during lymphopoiesis permits anergy (suppression of self-attack).
Naïve B cells each have one of millions of distinct surface antigen-specific receptors, yet have not encountered their specific, cognate antigen. With a life-span of only a few days, many B cells die without ever encountering their cognate antigen. Naïve B cells are activated when the BCR binds to its cognate antigen. This antigen-Ig binding must be coupled with a signal from a helper T cell in order to activate the B cell.
Once activated, B lymphocytes:
● differentiate into one of the B cell types (directly or through intermediate, germinal center reactions)
● plasma cells produce antibodies against the antigenic stimulus, or memory cells are primed for subsequent activation by the antigen
Types of B cell:
● B-1
● B-2
● Plasma B cells
● Memory B cells
After newly formed B cells exit generative sites in fetal liver or adult bone marrow they undergo selection events that may involve interactions with self or with external antigens. Selective events can influence the phenotype and functional characteristics of B cells. B cell receptor-mediated events also influence lymphoid organs localization as marginal zone B cells in the spleen, as follicular (B-2 cells), as well as B-1 cells in the peritoneal and pleural cavities. [] fluorescence micrograph spleen, fm high power in which T cells form periarteriolar lymphocyte sheath (PALS) (red) and B-2 cell follicles (green) []
{kind=link}
{kind=link}
B-1 cells are the first B cells produced in the fetus, and in adults are located primarily in the peritoneal and pleural cavities. B1 cells are believed to operate in the innate response to infection by viruses and bacteria, and usually show preferential responses to T cell-independent antigens. The diversity of B-1 lymphocytes is attributed to their recombinatorial recombination, in which there is a preferential recombination between D-proximal VH gene segments. B-1 lymphocytes express (polyspecific) IgM in greater quantities than they express IgG, and the ability of B1 cells to respond to isotype switch commitment factors such as interleukin-4 may be secondary to their production of IgM. B-1 cells express CD5, which binds to CD72 to mediate B cell-B cell interactions.
B-2 cells are conventional B lymphocytes that are produced postnatally (unlike fetal B-1 cells) and are replaced from the bone marrow.
Plasma B lymphocytes are committed to production of copious amounts of monoclonal antibodies.
Memory B lymphocytes are long-lived, stimulated B lymphocytes that are primed for rapid response to a repeated exposure of the priming antigen. Memory B cells are generated in lymphoid tissue after B cell activation/proliferation and reside in the bone marrow, lymph nodes, and spleen. High affinity surface immunoglobulins enable their activation by lower levels of cognate antigen than are naïve B cells.
NK cells are differentiated from killer T cells. NK, natural killer cells constitute a corps of circulating lymphocytes that are constitutively specialized to attack cancerous cells and virus infected cells. Preprogramming for target recognition, coupled with the absense of need for backup by a clone of identical cells, renders NK cells capable of rapid (innate) response to pathogens. NK attack involves the exocytosis of cytoplasmic granules containing perforin and granzymes. Perforin forms pores in the plasma membrane of attacked cells through which serine-protease granzymes enter, cleaving caspase precursors and triggering apoptosis.
Individuals inherit multiple, polymorphic genes for NK receptors, so the assemblage of NK receptors differs between individuals. NK cells carry two forms of surface receptors:
● killer inhibitory receptors (KIRs) transmit an inhibitory signal when they encounter class I MHC molecules on a cell surface. (By contrast, T cells only recognize antigens that are presented by a MHC molecule.)
● activating receptors, which activate the NK cell upon binding to a target cell
Viral infection often causes suppression of MHC expresion, leading to a reduction of inhibition of NKs by its killer inhibitory receptors. This double negative renders the virus infected cell a target for killing by NK cells.
"About 85% of peripheral B cells are phenotypically mature and display first-order exponential kinetics defined by a half-life of 5-6 weeks, whilst the remainder are short-lived with a life span of several days."[s]
[] tem plasma cell [] micrograph macrophage surrounded by normal plasma cells [] micrograph macrophage & plasma cells []
{kind=link}
{kind=link}
{kind=link}
▼ activation : BCRs : CDRs : helper T cells : life-span B cells : lymphopoiesis : naïve B cells : surface-immunoglobulins : surface receptors : VDJ recombination ▼
Tables Fc receptors Immune Cytokines Immunoglobulins Cell Adhesion Molecules Cell signaling Receptor Tyrosine Kinases (RTKs) Receptor Signal Transduction Second Messengers
▲ Top ▲
tags [Immunology][lymphocyte]
Labels: antibodies, B lymphocytes, BCR, humoral, immunoglobulins, plasma cells, VDJ recombination